Contrazione muscolare
La contrazione muscolare è il risultato di una serie di modificazioni intracellulari coordinate che porta a movimento della fibra muscolare e, di conseguenza, del muscolo stesso. La contrazione avviene in tutti i tipi di muscolo; è meglio rappresentata nel muscolo scheletrico, dove esiste una struttura metamerica (il sarcomero) dotata di particolarità morfologiche e funzionali.
|
Indice o
2.1 Contrazione nel muscolo
scheletrico o
2.2 Contrazione del muscolo
cardiaco |
Sarcomero
Il sarcomero è l'unità morfofunzionale e contrattile del muscolo striato scheletrico e cardiaco; i componenti peptidici principali sono:
- miosina (filamenti spessi disposti lungo il sarcomero)
- actina (filamenti sottili disposti lungo il sarcomero)
- altre proteine strutturali che permettono e favoriscono il mantenimento della struttura (ad esempio la distrofina).
Al microscopio si possono riconoscere alcune microstrutture, cioè:
Meccanismo di Contrazione
Contrazione nel muscolo scheletrico

Struttura macroscopica e microscopica del muscolo scheletrico
Il meccanismo di contrazione muscolare può essere riassunto nei seguenti
punti: - le linee Z si avvicinano -
- Lo stimolo alla contrazione, propagato attraverso le fibre nervose, giunge al bottone sinaptico dove determina il rilascio del neurotrasmettitore (solitamente acetilcolina).
- L'acetilcolina funge da ligando per alcuni canali del Na+ ligando dipendenti; causandone l'apertura, determina una depolarizzazione della membrana plasmatica della fibra.
- Questa prima minima depolarizzazione, causa a sua volta l'apertura di canali ionici per il Ca+, i canali diidropiridinici.
- L'apertura dei canali diidropiridinici causa, tramite cambiamenti conformazionali, l'apertura dei recettori rianodinici, canali ionici del Ca++ situati a livello del reticolo sarcoplasmatico.
- Dal reticolo sarcoplasmatico (compartimento cellulare in cui il Ca++ è solitamente sequestrato tramite apposite proteine) il Ca++ diffonde per tutta la cellula, raggiungendo ogni miofibrilla grazie ad un espediente morfologico della cellula, le triadi.
- Il Ca++ va a legarsi alla troponina, sulla subunità C; così facendo determina un cambiamento conformazionale che causa il trascinamento della subunità I, legata a sua volta alla tropomiosina. Questo movimento lascia libero il sito d'attacco per la miosina sull'actina, prima occupato dalla tropomiosina.

Cambiamenti conformazionali della miosina dovuti al legame con l'actina
La liberazione del predetto sito sull'actina dà il via alla fase di contrazione vera e propria. Lo scorrimento della miosina ed i suoi cambiamenti conformazionali, cause primarie della contrazione del muscolo, sono essenzialmente dovute alla diversa affinità della proteina per actina, ATP e ADP. Durante lo scorrimento le teste di miosina si legano a quelle di actina con una precisa angolazione di 45°. Nella fase di rilassamento, invece il procedimento sarà il contrario. Possiamo suddividere la contrazione e il rilassamento muscolare in tre fasi principali, ovvero la contrazione, il rilassamento ed infine la fase latente, fase che segue lo stimolo, ma nella quale non c’è risposta. Questo complesso sistema di reazioni chimiche determinerà lo scorrimento di un filamento sull’altro, e quindi la contrazione del sarcomero. A seguito della contrazione la troponina rilascia ioni Ca++ che tornano nel reticolo sarcoplasmatico.
Contrazione del muscolo cardiaco
differisce dal muscolo scheletrico per diversi aspetti. Le cellule del muscolo cardiaco mancano del sistema a "T" e non esiste un diretto accoppiamento fra la membrana plasmatica ed il reticolo sarcoplasmatico. Il potenziale d'azione cardiaco varia nella sua configurazione nelle diverse parti del cuore, ma in generale, mostra un plateau che dura parecchie centinaia di millisecondi e che è successivo alla fase di depolarizzazione rapida iniziale. La membrana plasmatica contiene molti canali del calcio tipo L, che si aprono durante questo plateau e permettono a Ca2 + di entrare nella cellula, anche se la quantità non è sufficiente ad attivare direttamente la macchina contrattile. Piuttosto, questo iniziale ingresso di Ca2 + permette a RyR (una molecola diversa da quella del muscolo scheletrico) di liberare Ca2 + da SR, dando origine a un'onda secondaria e di maggiori dimensioni di Ca2 + libero. Poiché i RyR del muscolo cardiaco sono essi stessi attivati da Ca2 + , l'onda di Ca2 + può essere considerata un evento di tipo rigenerativo "tutto o nulla". L'ingresso iniziale di Ca2 + che attiva questa situazione è dipendente dalla durata del potenziale d'azione e dal funzionamento dei canali del tipo L di membrana.
Contrazione nel muscolo liscio
Le differenze principali tra muscolo liscio e muscolo striato sono tre:
- nel muscolo liscio la contrazione può essere generata autonomamente o da ormoni (ad esempio epinefrina), oltre che da stimolo nervoso;
- nel muscolo liscio il Ca2+ liberato nel citosol proviene dall'ambiente extracellulare, e non dal reticolo sarcoplasmatico;
- il muscolo liscio non è dotato di sarcomeri o di altre unità funzionali. I filamenti di actina sono ancorati alla membrana plasmatica, generando una contrazione non organizzata delle cellula.
Il potenziale d'azione del muscolo liscio è un fenomeno piuttosto lento e vago se paragonato al comportamento più militaresco del muscolo scheletrico e di quello cardiaco, e si propaga attraverso il tessuto molto più lentamente e con incertezza. Nella maggior parte dei casi, il potenziale d'azione è generato da canali del calcio del tipo L piuttosto che da canali del sodio voltaggio-dipendenti (questa rappresenta una via piuttosto importante per l'ingresso di Ca2 + ). In aggiunta molte cellule del muscolo liscio possiedono canali cationici attivati da ligandi, che permettono l'ingresso di Ca2 + quando vengono attivati dai neurotrasmettitori. Tra questi, i recettori meglio caratterizzati sono i quelli del tipo P2x, che sono attivati dall'ATP rilasciato dalle terminazioni nervose del sistema autonomo. Anche le cellule del muscolo liscio accumulano Ca2 + nel reticolo endoplasmatico, da cui può essere rilasciato quando IP3R viene attivato da IP3. IP3 viene attivato dalla generazione di molti tipi di recettori accoppiati alle proteine G. Quindi, a differenza della muscolatura scheletrica e liscia, il rilascio di Ca2 + e la conseguente contrazione possono verificarsi nel muscolo liscio quando questi recettori vengono attivati senza che ci sia alcun coinvolgimento di eventi elettrici e di ingresso di Ca2 + attraverso la membrana plasmatica. Il sistema contrattile della muscolatra liscia viene attivato quando la catena leggera della miosina (MLC) viene fosforilata e può così staccarsi dai filamenti di actina. Questa fosforilazione viene catalizzata da una chinasi, la chinasi della miosina a catena leggera (MLCK), che viene attivata quando si lega alla Ca2 + -calmodulina. Un secondo enzima, la miosina fosfatasi, defosforila e promuove il rilasciamento. Entrambi gli enzimi sono regolati da nucleotidi ciclici (cAMP
e cGMP), e molti farmaci che causano la contrazione o il rilasciamento della muscolatura liscia agiscono in questo modo mediante attivazione di recettori accoppiati a proteine G o alle guanilato ciclasi.Voci correlate
Actina

L'actina è una proteina di forma globulare, con un diametro di circa 7 nm, dal peso di 43 kDa (leggi: kilo Dalton) e costituisce una porzione abbondante (5-10%) di tutte le proteine delle cellule eucariote.

La più alta presenza di actina si verifica nelle cellule del tessuto muscolare (circa 20% delle proteine totali), dove è fondamentale per il processo di contrazione.
Ogni singola subunità di actina (detta actina G, cioè globulare) si può legare ad altre due subunità, formando così un polimero lineare. Due polimeri lineari avvolti tra di loro danno origine ad un microfilamento, uno dei tre tipi fondamentali di filamenti che compongono il citoscheletro.

Un microfilamento; in due toni di azzurro le subunità di actina di un semifilamento e in due toni di verde le subunità del secondo semifilamento. In colori misti è evidenziata una singola subunità di actina.
|
Indice ·
1 Assemblaggio e disassemblaggio |
Assemblaggio e disassemblaggio
L'assemblaggio dei filamenti di actina è dipendente dalle condizioni
dell'ambiente interno alla cellula, il citosol,
ed in particolare dagli ioni
presenti, dalle loro concentrazioni e dal legame con l'ATP, l'adenosinatrifosfato,
un'importante molecola energetica.
La polimerizzazione è infatti favorita da una concentrazione salina pari a
quella corporea; inoltre, i monomeri di actina legati
ad ATP tendono ad aggiungersi e a restare attaccati al filamento con maggiore
facilità rispetto ai monomeri in cui l'ATP è
diventata ADP, l'adenosindifosfato.
Le attuali conoscenze sulla struttura dei microfilamenti sono basate su
fotografie al microscopio elettronico dell'actina F e su
modelli derivanti da studi ai raggi X di gel orientati di actina F. Tali modelli
evidenziano che il polimero di actina è una catena doppia, in cui ciascuna subunità interagisce con altre quattro. Le varie subunità, disposte in fila all'interno di una catena, suono
ruotate di circa 167° l'una rispetto all'altra, dando così l'aspetto di
"elica con gobbe" al microfilamento. Ciascuna subunità
presenta inoltre lo stesso orientamento testa-coda, in modo che la fibra intera
abbia una precisa polarità. La presenza di un capo del filamento caratterizzato
da actina legata ad ATP identifica l'estremità "più", mentre l'altro
capo, dove l'actina è legata a ADP costituisce l'estremità "meno" del
filamento. A causa di tale differenza strutturale fra le due estremità, la
velocità di aggiunta di subunità è maggiore
all'estremità "più" che di conseguenza cresce in lunghezza più
rapidamente dell'estremità "meno".
Organizzazione dei filamenti e proteine associate
I singoli microfilamenti vengono generalmente organizzati e integrati dalle proteine leganti l'actina (ABP, Actine Bindig Proteins). Questa famiglia di proteine si può suddividere in sottoclassi a seconda del tipo di interazione che intrattiene con l'actina:
- proteine che favoriscono la formazione dei filamenti: profilina ( si lega ai singoli monomeri di actina);
- proteine che ostacolano la formazione dei filamenti: severina e timosina (si legano anch'esse ai singoli monomeri di actina), gelsolina che taglia il filamento e si lega all'estremità "più" appena formata, katanina;
- proteine che organizzano i microfilamenti in fasci paralleli: fimbrina, α-actinina, tropomiosina, fodrina, villina;
- proteine che organizzano i microfilamenti in reticoli: filamina;
- proteine che collegano i microfilamenti alla membrana plasmatica: spettrina, distrofina, vinculina, talina, tensina;
- proteine che si muovono sui microfilamenti: miosina.
Esiste anche una distinzione in funzione al fatto che le ABP siano legate
con G-Actina (forma globulare actina+atp) o F-Actina
(filamentosa). Un esempio delle ABP capaci di legarsi con le molecole di
G-Actina sono la B-Timosina e
Funzione
La singola subunità di actina non espleta direttamente una funzione, ma lo fa tramite la struttura che forma, il già citato microfilamento. Nella cellula sono presenti moltissimi microfilamenti e questi assolvono a diverse funzioni:
- forniscono sostegno e forma alla cellula, collegandosi con gli organelli interni e con la membrana plasmatica;
- permettono una parte del trasporto interno di vescicole;
- i sarcomeri, le unità contrattili delle cellule muscolari sono formati da actina, oltre che da miosina ed altre proteine;
- costituiscono la base su cui si formano le principali estroflessioni cellulari, quali microvilli, stereociglia, filopodi, lamellipodi, pseudopodi;
- sono coinvolti nell'adesione tra cellule e tra una cellula ed un substrato.
Miosina
Le miosine sono una vasta famiglia di proteine motrici, rintracciabili in cellule eucariotiche, responsabili del movimento basato sui filamenti di actina.
|
Indice ·
4
Note |
Struttura e funzione

Modello del funzionamento della contrazione muscolare.

Particolare della regione globulare della miosina
La tipica struttura della miosina consiste essenzialmente in due domini, definiti testa e coda:
- la testa è una regione globulare deputata al legame con i filamenti di actina, oltre che al sito di idrolisi dell'ATP. Questa reazione chimica permette il movimento della miosina verso l'estremità + del filamento di actina, tramite cambiamenti conformazionali dovuti alla diversa affinità della proteina per le molecole che può legare (ATP, actina, ADP in ordine decrescente di affinità).
- la coda è un dominio di struttura allungata che generalmente media le interazioni con molecole trasportatrici e subunità di miosina.
Miosina II
Miosina VI
Differenze tra miosine

Albero filogenetico delle miosine
L'analisi delle sequenze amminoacidiche mostra una elevata conservazione dei residui della testa ed un'alta variabilità delle code. Ciò può essere spiegato dal fatto che, se le code delle diverse miosine devono essere in grado di legare un elevato numero di molecole terze, le teste sono deputate esclusivamente al movimento ed il meccanismo motore è dunque rimasto immutato lungo il corso dell'evoluzione.
Le differenze riscontrate tra le diverse forme di miosina sono legate anche
a notevoli differenze nella velocità del movimento lungo i filamenti di actina. In tutte le
miosine, infatti, l'idrolisi di ATP ed il successivo rilascio di un gruppo
fosfato, generano una flessione presso la testa, con la proiezione in
avanti della regione definita del collo. Le forme che presentano un collo
più lungo, tuttavia, sono in grado di coprire una maggiore distanza a parità di
idrolisi di ATP (nello stesso modo attraverso cui persone con gambe più lunghe
sono in grado di coprire una maggior distanza con un passo).
Geni nell'uomo
I geni per i componenti della miosina sono divisi in diverse famiglie. Non tutti i geni riportati sono effettivamente espressi.
- Famiglia I: MYO1A, MYO1B, MYO1C, MYO1D, MYO1E, MYO1F, MYO1G, MYO1H;
- Famiglia III: MYO3A, MYO3B;
- Famiglia V: MYO5A, MYO5B, MYO5C;
- Famiglia VI: MYO6;
- Famiglia VII: MYO7A, MYO7B;
- Famiglia IX: MYO9A, MYO9B, MYO10;
- Famiglia XV: MYO15A;
- Famiglia XVIII: MYO18A, MYO18B;
- Catene pesanti: MYH1, MYH2, MYH3, MYH4, MYH6, MYH7, MYH7B, MYH8, MYH9, MYH10, MYH11, MYH13, MYH14, MYH15, MYH16;
- Catene leggere: MYL1, MYL2, MYL3, MYL4, MYL5, MYL6, MYL6B, MYL7, MYL9, MYLIP, MYLK, MYLK2, MYLL1.
|
fase 1 |
fase 2 |
fase 3 |
fase 4 |




|
|
|
|
|
|
Contrazione Muscolare |
contrazione muscolare, |
Contrazione muscolare |
contrazione muscolare |
|
|
|
|
|
|
(contrazione
muscolare |
contrazione muscolare |
Contrazione nel muscolo |
quanto
la loro contrazione |
|
|
|
|
|
|
alla
contrazione muscolare |
CONTRAZIONE MUSCOLARE |
Modello
contrazione muscolo |
|
|
|
|
|
|
|
La
contrazione muscolare) |
la
contrazione muscolare. |
contrazione muscolare |
Poichè ciascun muscolo |















Contrazione
muscolare
Miosina, ATP e
contrazione muscolare
Il muscolo
scheletrico può essere paragonato ad un motore, in grado di convertire
l'energia chimica, fornita dall'ATP, in energia meccanica,
agendo sul sistema di leve
scheletriche con un buon rendimento (non più del 30-50% di questa energia
viene dissipata come calore). Il risultato di questa reazione endoergonica è la contrazione muscolare.
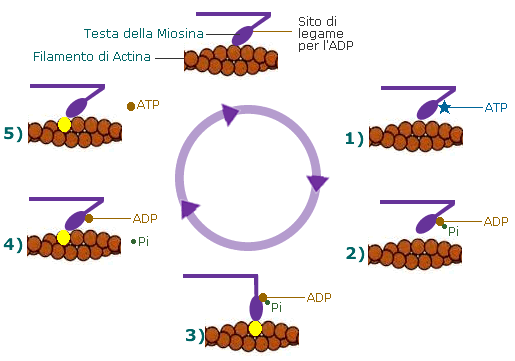
Ogni
molecola della miosina presenta due siti di legame, uno per una molecola di ATP
ed uno per l'actina. La sua attività
ATPasica le consente di idrolizzare l'ATP ad ADP
+ fosfato inorganico e di utilizzare l'energia così sviluppata per generare il
movimento. Il tutto avviene con un ciclo di eventi molecolari:
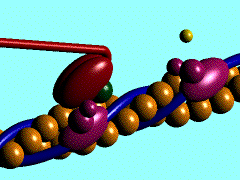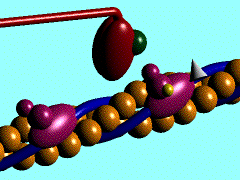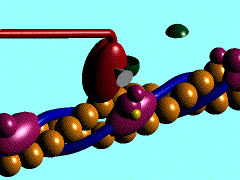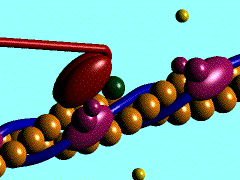
1)
L'ancoraggio dell'ATP nella specifica sede di legame sulla testa
della miosina porta al distacco di quest'ultima dalla molecola di G-Actina
2) L'ATP,
legata alla testa miosinica, viene idrolizzata ad ADP
e fosfato inorganico (Pi); entrambi i prodotti rimangono ancorati in
questa sede; per consentire tale reazione sembra necessaria la presenza di magnesio.
3)
L'energia liberata dall'idrolisi dell'ATP induce una rotazione della testa
della miosina che, caricandosi di energia potenziale, si lega debolmente ad una
molecola di G-actina con un angolo di 90°.
4) Il
rilascio del fosfato inorganico provoca un cambiamento conformazionale nella
testa della miosina, generando il cosiddetto colpo di frusta. La fune (il
filamento di actina) viene così tirata verso il centro del sarcomero,
vale a dire verso
5) La testa
della miosina rilascia anche la molecola di ADP e rimane strettamente ancorata
all'actina, in uno stato di rigor che dura soltanto pochi istanti, prima che il
ciclo ricominci con l'ennesimo legame miosina-ATP.
|
|
|
||
|
Il
meccanismo di CONTRAZIONE MUSCOLARE da: www.sci.sdsu.edu/movies/actin_myosin.html |
|||
|
|
La rinite allergica è una patologia di frequente riscontro,
che consiste nell'infiammazione della mucosa nasale provocata
dall'esposizione a determinate sostanze. Nei soli Stati Uniti, gli individui
che soffrono di rinite allergica sono oltre 50 milioni. |
||